December 10, 2019
Background
Animal cognition is a multidisciplinary field of study that spans fields such as biology, evolution, psychology, and neurology. The areas pertaining to animal cognition deals with how animals process information brought in by their senses and how they then act on that information. Cognitive studies do not imply the existence of consciousness in the animals, but instead seek only to assess how animals process new information, thus shaping their specific behaviors. The understanding of specific behaviors can be applied to animal distributions, ecosystem ecologies, and studying inter and intraspecies interactions. Sexual interactions and strategies can ultimately shape entire species in distribution and spread.
Why do animals vary in their cognitive abilities? Classic comparative approaches allow researchers to model how human cognition evolved from a vertebrate ancestor (comparative cognition), or how similar environments may select for convergence in cognitive traits across species (cognitive ecology). However, such approaches cannot explain the prevalence of cognitive variation within a species. A recent focus on individual differences in behavior has prompted a parallel interest in how those differences might relate to cognitive variation (Boogert et al. 2018). Species with alternative reproductive tactics offer a unique opportunity to understand how individual differences in behavior relate to cognitive variation. Here, I propose to test how discrete variation in reproductive behavior relates to anxiety/boldness and spatial cognition.
I will be working to understand how differences in male sexual tactics can lead to cognitive and behavior variation between male, poeciliid fish. I will be using Xiphophorus nigrensis, the El Abra Swordtail fish, as the study subject. The males of this species have three separate morphologies with well-documented differences in sexual behavior. The small male phenotype adheres to a coercive mating tactic of sneak copulation, and the large male phenotype a courting technique through dance and display to successfully copulate. The third male morphology is the intermediate male, and this male type exhibits both coercive and courting tactics to achieve a successful copulation.
Although the bright coloration of large and intermediate males increases female attention, there is an energetic cost associated with such a display and with avoiding predators who are also attracted to conspicuous displays. On the converse, the small male has to avoid being chased by the larger males as well as encounter many unsuccessful copulations with the females. Because each of these phenotypes have specific trade-offs influencing their mating strategies, it is reasonable to think they have different cognitive variation and functions in regard to their mating strategies and counter strategies.
Objectives
I aim to find if differing sexual strategies and behaviors have an effect on cognitive abilities. Studies on the cognitive flexibility and variation between females and males of poeciliid fish have been conducted (Lucon-Xiccato and Bisazza 2016), but discrete differences in cognition only in the male population have not been studied. First, I hypothesize that the intermediate male type will demonstrate the best performance in the detour reaching assay, and that this performance is due to their ability to span two cognitively separate sexual techniques. This varying behavioral tendency could mean they have to be more adept to know when to use each tactic, and thus, this behavioral variability likely relates to cognitive flexibility. Secondly, I hypothesize that the less anxious and more exploration-oriented males will demonstrate better performances in the detour reaching assay due to their performance in a novel environment like scototaxis. Finally, I hypothesize that the small males will perform better in the route learning assay, due to their tendencies to seek safe areas away from predators and large males, and they are more likely to seek out shoals for protection.
Methods
A total of 48 male fish (12 from each size class) from a semi-wild environment at the University of Texas Brackenridge Field Laboratory will be used for this experiment. The most promising results in cognitive analyses have been from conducting a diverse array of repeated cognitive performance tests (Boogert et al. 2018). Because of this, each of the male fish will be placed through two cognitive assays and three behavioral assays over the course of a week, each assay testing a different aspect of cognitive ability. For this report, however, only data from the scototaxis (anxious/boldness test), detour-reaching, and route learning will be analyzed. Prior to each assay, the males are isolated for a day to control prior experience. The males will first be placed through an assay to assess their reactions to a stressful environment: scototaxis. By making a tank half white (anxiety inducing environment for fish) and half black (a more comfortable environment), this assay will be used to test exploration tendencies and anxiety of the males (Schumm et al., in review 2019). The fourth test of the week will be a detour-reaching assay to assess the males’ ability to inhibit basic motor controls and reach a reward behind a transparent barrier by travelling around the edges of the barrier. Each fish will be placed through this assay one time to test for learning ability and subsequent performance increases between each trial. Finally, the males will be placed through a route-learning assay to assess spatial cognition and learning capabilities. There are two sections of the maze that each fish will have to navigate through, each section divided by a maze-like barrier with a false entry port on one side. Visual cues will be placed by the entrance that leads to the next section, so placing each male through the route learning assay five separate times will test if they can learn how to navigate and successfully reach the reward. The reward at the end is a female, and behind the female is a more natural section of the tank with a shoal visible from an adjacent tank. These rewards can be seen through transparent windows in the middle that span the length of the tank. The coloration throughout the sections of this assay goes from white to light brown to green—from anxiety inducing coloration to more comfortable coloration.
Significance
Because studies including both route learning and detour reaching assays have been used to assess cognitive flexibility differences between poeciliid sexes (Lucon-Xiccato and Bisazza 2016), it is important to study this difference between individual males in poeciliid fish. By looking at individual cognitive traits between male types, we hope to find significant results that indicate the existence of intra-sexual selection of cognition. Understanding the cognitive flexibility and variation of poeciliid males on the individual level will give more insight into how and why these variations were selected for between individual males.
While some have attempted to link animal personality and cognition together, there are practical issues in testing this due to the complexities of personality. However, the analysis of this data can hopefully lead towards greater understandings of behavioral ecology and the adaptive selection of these traits in poeciliid fish (Griffin 2015). Individual cognitive variation has remained relatively unstudied, but there is increasing evidence for its ecological importance (Sih; Wolf and Weissing 2012). Furthermore, the discrete differences in anxiety and boldness tendencies of males can shape cognition selection. Other studies have shown that male sexual performance in competitive environments is linked to spatial cognition (Smith 2015), so we hope to find indication that the performance of X. nigrensis in these tests is linked to anxiety/boldness tendencies and spatial learning performance.
Preliminary Results and Future Steps:
At the completion of the Fall 2019 semester, we have successfully run 3 fish, with the hopes of completing assays for 6 fish per week in the coming months. Scoring for route learning was done manually, and the data points used for the following graph were the total times taken to reach the reward for each of the five trials the three fish went through.
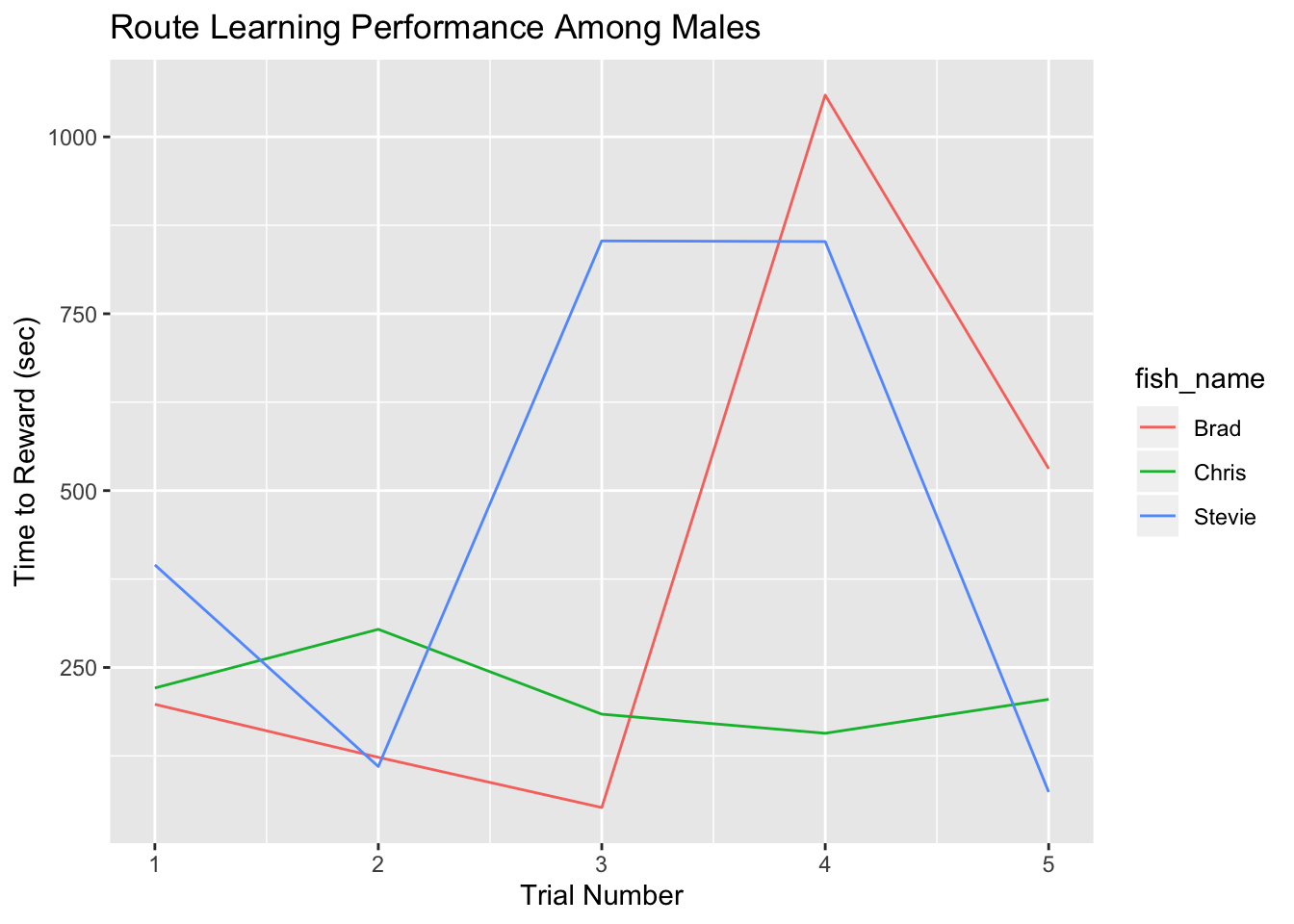
Although significant conclusions cannot be inferred from this data, there is a trend of fish Brad (large male) and Stevie (intermediate male) who completed the assay quicker on the fifth trial. As for Chris (small male), there wasn’t a learning trend that can be deduced from his performance in the trials. All three fish actively participated in the trials, even though the times for completion for Brad and Stevie were very high on the fourth trials. The first choice of route that each fish made was recorded as well, and this could also be a valuable metric for determining whether the males are learning from previous exposures. With the detour and scototaxis assays being scored, these data will be included in future analysis to gain a more holistic insight into what shapes individual cognition.
References
Boogert, NJ, Madden, JR, Morand-Ferron, J, Thornton, A. (2018) Measuring and understanding individual differences in cognition. Phil. Trans. R. Soc. B 373: 20170280. http://dx.doi.org/10.1098/rstb.2017.0280
Lucon-Xiccato, T, A Bisazza. (2016) Sex differences in spatial abilities and cognitive flexibility in the guppy. Animal Behavior. 123 (2017): 53-60.
Schumm, M, S Pineau, P Mackey, V Sakthivelnathan, M Cummings, M Ramsey. (2019) Variation in cognitive and noncognitive components of a detour maze: Poeciliid fish vary by species, sex and context. (unpublished)
Shettleworth, SJ. (2000) Animal cognition and animal behavior. Animal Behavior. (2001) 61: 277-286. Smith C, Philips A, Reichard M. 2015 Cognitive ability is heritable and predicts the success of an alternative mating tactic. Proc. R. Soc. B 282: 20151046. http://dx.doi.org/10.1098/rspb.2015.1046 Sih, A, MD Giudice. (2012) Linking behavioural syndromes and cognition: a behavioural ecology perspective. Royal Society Publishing. 367: 2762-2772.
Wolf, M, J Weissing. (2012) Animal personalities: consequences for ecology and evolution. Cell Press: Trends in Ecology and Evolution. 27(8): 452-461.